- Tiếng Việt
- English
Con đường sinh tổng hợp của artemisinin (artemisinin biosynthesis)
Artemisinin là hợp chất quý thuộc nhóm Sesquiterpene endoperoxidized và được tìm thấy trong cây thanh cao hoa vàng. Hoạt chất này đem lại nhiều tác dụng trong việc chăm sóc sức khỏe con người. Chính vì thế quá trình sinh tổng hợp artemisinin đã được nghiên cứu trong nhiều năm vì lợi ích khoa học và cho các ứng dụng sản xuất tiềm năng để đáp ứng nhu cầu thị trường.
1. Ý nghĩa của quá trình sinh tổng hợp artemisinin (Role of artemisinin biosynthesis)
Bệnh sốt rét là một trong những nguyên nhân hàng đầu gây ra dịch bệnh truyền nhiễm và gây tử vong cho nhiều người trên toàn thế giới. Theo số lượng ước tính, mỗi năm có khoảng 400 triệu ca nhiễm bệnh sốt rét và 1 đến 3 triệu ca tử vong.
Bệnh xảy ra nhiễm ký sinh trùng sốt rét qua vật chủ trung gian là muỗi và để lại những hậu quả, biến chứng vô cùng nguy hiểm. Trẻ nhỏ và phụ nữ mang thai là những đối tượng dễ bị mắc bệnh này nhất.
Và thật may mắn là năm 1971, các nhà khoa học Trung Quốc đã phát hiện ra rằng hoạt chất artemisinin có trong cây thanh cao hoa vàng có thể điều trị được căn bệnh này.
Nhiều dẫn xuất của artemisinin cũng đã được tổng hợp và thử nghiệm chống lại ký sinh trùng sốt rét. Chẳng hạn như trioxolane OZ 277, đã được đưa vào nghiên cứu lâm sàng trong năm 2004. Hợp chất trioxolane này được dùng bằng đường uống và có hiệu lực cao, nhưng quá trình nghiên cứu thuốc này đã bị tạm ngừng trước các thử nghiệm lâm sàng giai đoạn 3 vì tính không ổn định của nó trong máu.
Vì vậy, nhu cầu cần thêm thuốc chống sốt rét vẫn vô cùng cấp thiết. Các thuốc là dẫn xuất của artemisinin thường có thời gian bán hủy ngắn và được sử dụng tốt nhất khi kết hợp với các thuốc chống sốt rét khác như lumefantrine, sulfadoxime/pyrimethamine trong các liệu pháp kết hợp dựa trên artemisinin (ACT).
Ước tính trên toàn thế giới mỗi năm cần có 400 - 600 triệu liều điều trị artemisinin mỗi năm, trong khi hiện tại chúng ta mới chỉ đáp ứng được chưa đến 100 triệu liều mỗi năm. Do đó, có thể thấy được rằng nhu cầu sử dụng artemisinin trong điều trị bệnh và sử dụng artemisinin để tổng hợp các dẫn xuất khác đang rơi vào tình trạng thiếu hụt trầm trọng.
Hơn thế nữa, hiện trên thị trường artemisinin chỉ được sản xuất từ cây thanh cao hoa vàng (A. annua) và loại thảo dược này chỉ chứa một lượng nhỏ artemisinin ở khoảng 0,01% đến 0,8%.
Không những vậy, sản lượng của artemisinin còn phụ thuộc vào nhiều điều kiện thời tiết trong quá trình cây thanh cao hoa vàng sinh trưởng và phát triển như: nhiệt độ, độ ẩm và loại đất.
Do đó, các nhà khoa học hiện đang rất nỗ lực để tăng sản xuất, tổng hợp nên hợp chất artemisinin. Và sinh tổng hợp chính là một trong giải pháp để khắc phục tình trạng này.

Quá trình sinh tổng hợp artemisinin đem lại nhiều lợi ích
2. Con đường sinh tổng hợp của artemisinin
Theo các nhà nghiên cứu, quá trình sinh tổng hợp artemisinin thường diễn ra qua 2 giai đoạn:
- Sản xuất các tiền chất isopren bao gồm geranyl pyrophosphate (GPP), farnesyl pyrophosphate (FPP) và geranyl terpen.
- Sự hình thành artemisinin từ các vòng terpen.
Sự hình thành của FPP, GPP và geranyl terpen
Trong nhiều thập kỷ, con đường mevanolate (MVA) đã được coi là con đường duy nhất cho quá trình sinh tổng hợp terpen, được phát hiện lần đầu bởi Bloch và Lynen vào năm 1958.
Để tổng hợp FPP thì cần có hai tiền chất isoprenoid, isopentenyl diphosphate (IPP). , được sinh tổng hợp thông qua mevalonate có nguồn gốc từ acetyl-CoA.
Tuy nhiên, một con đường sinh tổng hợp khác đã được làm sáng tỏ gần đây. Con đường plastidic non-MVA, hoặc con đường deoxyxylulose phosphate (DXP)/methylerythritol phosphate (MEP), bắt nguồn từ glyceric acid-3-phosphate và pyruvate.
Nghiên cứu ban đầu chỉ ra rằng isopentenyl diphosphate (IPP), chất khởi đầu của quá trình -mevalonate đóng vai trò là tiền chất của farnesyl diphosphate (FPP). Trong khi đó, chu trình non-mevalonate cung cấp tiền chất của geranyl diphosphate (GPP) để tổng hợp sesquiterpen.
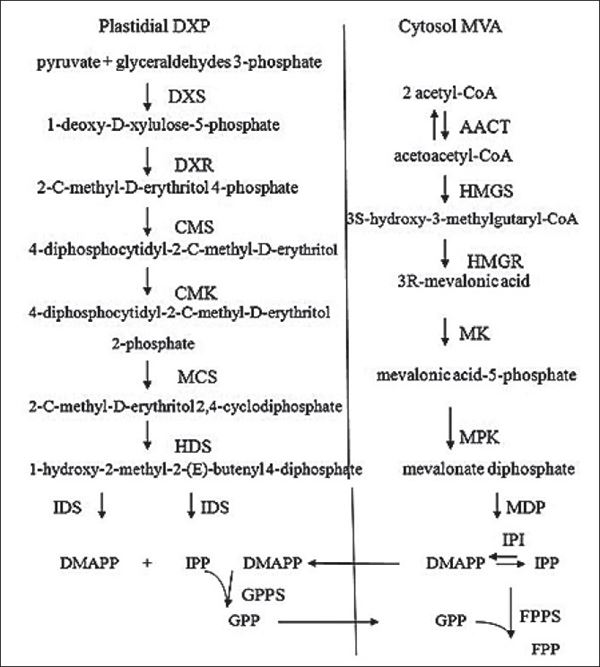
Quá trình tạo thành FPP, GPP
Sự hình thành của FPP được xúc tác bởi một loạt các enzyme, trong quá trình MVA có sự tham gia của:
- ATOT (acetoacetyl-CoA thiolase).
- HMGS (3-hydroxyl-3-methyglutaryl CoA synthase).
- HMGR (3-hydroxyl-3-methyglutaryl CoA reductase).
- MK (mevalonate -phosphate kinase).
- MPD (mevalonate pyrophosphate decarboxylase).
- IPI (IPP isomerase).
Đối với các con đường non – mevanolate, có sự tham gia của các enzyme như:
- DXS (1-deoxy-D-xylulose-5-phosphate synthase).
- DXR (1-deoxy-D-xylulose-5-phosphate reductoisomerase).
- CMS (4-diphosphocytidyl-2C- methyl-D-erythritol 4-phosphate synthase).
- CMK (4-diphosphocytidyl-2-C-methyl-D-erythritol kinase).
- MCS (2-C-methyl-D- erythritol 2,4-cyclodiphosphate synthase).
- HDS ( 1-hydroxy-2-methyl-2- (E) -butenyl 4-diphosphate synthase).
- IDS (IPP / DMAPP synthase).
HMGR có ý nghĩa bằng cách xúc tác HMG-CoA thành 3R-mevalonic acid và được coi là enzyme giới hạn đầu tiên trong con đường MVA, để đảm bảo cho quá trình 3R-mevalonic acid là một không thể đảo ngược.
Các sản phẩm nhân bản genome hoặc cDNA của HMGR trong cà chua, khoai tây, Arabidopsis thaliana, v.v. đã được báo cáo trong những ngày gần đây, và trong A. annua cho đến năm 1995, được nhân bản bởi Kang. Các gen của HMGR và FPPS đã được đồng thể hiện trong các cây thanh cao hoa vàng biến đổi gen, tạo ra sự gia tăng hàm lượng gấp 2,32 lần của artemisinin.
Các thí nghiệm điều chỉnh hoạt động của HMGR để tăng sinh tổng hợp artemisinin thông qua việc cung cấp HMG-CoA có nhãn ngoại sinh, và chứng minh rằng các chất điều hòa sinh trưởng thực vật, IAA và GA3, đã tăng cường tích lũy artemisinin bằng cách tăng hoạt động của HMGR.
DXS đã xúc tác bước đầu tiên trong quá trình n-MVA, thúc đẩy sự hình thành DXP từ pyruvate và glyceric acid-3-phosphate. DXR là một enzyme khác tham gia quá trình n-MVA với mục tiêu điều chỉnh sinh tổng hợp terpene.
Việc cung cấp chất ức chế cạnh tranh của DXR như fosmidomycin có thể đã ức chế 14,2% sự tích lũy artemisinin.
Do đó mà khi ngưng tụ tiếp DMAPP và IPP có thể tạo thành tiền chất prenyl diphosphate, GPP và FPP và quá trình này thường được xúc tác bởi prenyltransferase: GPP synthase (GPPS) và FPP synthase (FPPS).
Sự hình thành artemisinin từ các vòng terpen và các enzyme có liên quan
Sesquiterpene synthases
Một số cDNA từ A. annua mã hóa các synthase sesquiterpene khác nhau đã được tiết lộ. Trước hết, epi-cedol synthase (ECS), sau đó là nhân bản di truyền của amorpha-4,11-diene synthase (ADS) và caryophyllene synthase (CS).
Sau đó, các nhà khoa học đã nhân bản một (E)-farnesene synthase (FS) có trong cây thanh cao hoa vàng.
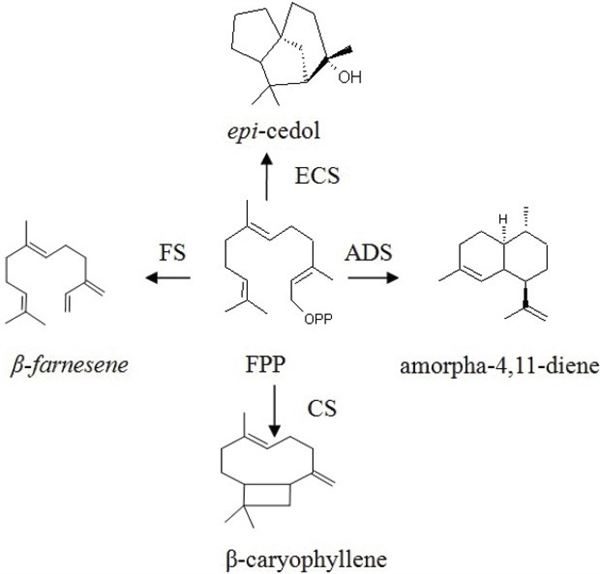
Serquiterpen synthase
Tất cả các synthase sesquiterpene đã đề cập ở trên đã được nhân bản từ lá của cây thanh cao hoa vàng. Tuy nhiên, để nghiên cứu quá trình sinh tổng hợp của Sesquiterpenes, nhà khoa học Bertea và các cộng sự đã phân lập tất cả RNA từ lông tuyến (glandular trichomes) và sử dụng chúng để xây dựng cDNA plasmid.
Bước đầu tiên để giúp tăng hiệu suất của quá trình sinh tổng hợp artemisinin cần phải tác động tới chu kỳ của FPP với sự tham gia của ADS. Do đó, việc điều chỉnh tăng chất vô định hình 4,11- diene synthase được coi là một chiến lược khả thi để tăng sản xuất artemisinin từ cây thanh cao hoa vàng.
Gen ADS đã được nhân bản ở cây thanh cao hoa vàng, dẫn đến hàm lượng artemisinin tăng gấp 2,3 lần so với loại cây này khi chưa biến đổi gen.
Cytochrom P450
Enzyme Cytochrom P450 (CYP/P450) cùng với hoạt động của enzyme dehydrogenase và reductase được cho là tham gia vào quá trình oxy hóa C12 trong amorpha-4,11-diene.
Hơn nữa, một bản sao cDNA mã hóa một cytochrom P450 được chỉ định CYP71AV1 đã được đặc trưng bởi sự biểu hiện trong Saccharomyces cerevisiae và được hiển thị để xúc tác quá trình oxy hóa của amorpha-4,11-diene, alcohol artemisinic và aldehyde artemisinic.
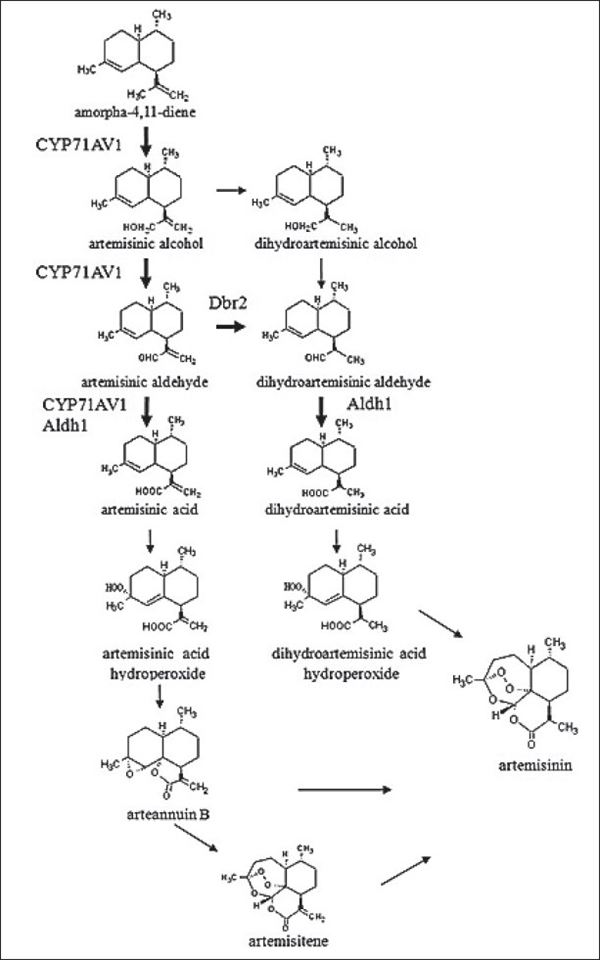
Sự tham gia của cytochrom P450
Artemisinic aldehyde Δ11 (13) reductase
Liên kết đôi ∆11 (13) trong phân tử amorpha-4,11-diene bị khử trong quá trình sinh tổng hợp artemisinin, để tạo ra gốc –CHO trong phân tử artemisinic aldehyde.
Gần đây, một gen Dbr2 tương ứng đã được nhân bản và được chứng minh là tương đối đặc hiệu với aldehyd artemisinic và một số hoạt động trên các hợp chất α,β - cacbonyl chưa bão hòa. Biểu hiện trong nấm men có gen Dbr2 và các enzyme khác của con đường artemisinin trong nấm men dẫn đến sự tích tụ acid dihydroartemisinic.
Một số cây thanh cao hoa vàng có nồng độ artemisinin cao thường có tỷ lệ acid dihydroartemisinic/acid artemisinic, trong đó, quá trình khử liên kết đôi có thể hạn chế tích lũy artemisinin.
Aldehyd dehydrogenase
Như đã đề cập ở trên, cytochrom P450 CYP71AV1 đã được chứng minh là có khả năng oxy hóa rượu artemisinic thành aldehyd và acid tương ứng.
Một cDNA có chiều dài đầy đủ mã hóa một homolog aldehyd dehydrogenase đã được phân lập và được đặt tên là Aldh1, có khả năng xúc tác cho aldehyd artemisinic oxy hóa.
Ngoài ra, biểu hiện cao nhất của Aldh1 cây thanh cao hoa vàng được quan sát thấy ở lông tuyến trong nụ hoa với mức độ vừa phải và biểu hiện thấp ở lá.
Các chất trung gian và enzyme khác
Toàn bộ con đường sinh tổng hợp từ amorpha-4,11-diene đến artemisinin hiện vẫn đang trong quá trình nghiên cứu. Vấn đề chủ yếu được các nhà khoa học quan tâm là về các bước mới nhất trong sinh tổng hợp artemisinin rằng liệu axit artemisinic hoặc dihydroartemisinic đóng vai trò là tiền chất sau này.
Axit artemisinic là tiền chất phổ biến của arteannuin B và artemisinin. Các nhà khoa học đã tinh chế và mô tả một loại enzyme liên quan đến chuyển hóa sinh hóa của arteannuin B thành artemisinin từ cây thanh cao hoa vàng (A. annua).
Tuy nhiên, gen của enzyme này đã không được báo cáo. Bằng chứng ở trên cho thấy khả năng sinh tổng hợp artemisinin từ axit artemisinic.
Hơn nữa, một cDNA (apod1) có độ dài đầy đủ mã hóa một peroxidase đã được phân lập từ A. annua, trong đó ưu tiên chuyển đổi sinh học của axit artemisinic thành artemisinin trong chiết xuất A. annua một cách gián tiếp nhưng không bị oxy hóa với axit artemisinic.
Gần đây, Brown và cộng sự đã báo cáo rằng axit artemisinic đã được chuyển đổi in vivo, có thể không có enzyme thành một số hợp chất bao gồm arteannuin B, nhưng không phải là artemisinin.
Ngoài ra, trong các nghiên cứu in vivo, acid dihydroartemisinic trải qua quá trình oxy hóa sắc tố thực vật nhanh chóng, sau đó là quá trình oxy hóa tự phát tiếp theo để tạo thành artemisinin.
Axit dihydroartemisinic có thể hoạt động như một chất khử oxy oxy đơn để sản xuất acid dihydroartemisinic hydroperoxide, sau này được chuyển thành artemisinin.
Đáng chú ý, có lẽ là bước tự phát ít nhất trong quá trình oxy hóa acid dihydroartemisinic, hydroperoxid hóa phụ thuộc oxy đầu tiên và tạo ra sự phân tách vòng rất gợi nhớ đến các phản ứng enzyme khác, như sự phân tách các hợp chất của carotene.
Vấn đề mà các nhà khoa học quan tâm lúc này là nếu tính đến các ý tưởng trên, liệu sinh tổng hợp artemisinin từ acid dihydroartemisinic có liên quan đến enzyme hay không là một câu hỏi mở và đáng để nghiên cứu thêm.
Mong rằng qua những thông tin mà bài viết đã chia sẻ đã giúp các bạn có thêm nhiều thông tin về sinh tổng hợp artemisinin. Chúc bạn có nhiều sức khỏe và thành công hơn nữa.


Sản phẩm










![]()
Viết bình luận